Neuroscience
Arturo Tozzi
Former Center for Nonlinear Science, Department of Physics, University of North Texas, Denton, Texas, USA
Former Computationally Intelligent Systems and Signals, University of Manitoba, Winnipeg, Canada
ASL Napoli 1 Centro, Distretto 27, Naples, Italy

For years, I have published across diverse academic journals and disciplines, including mathematics, physics, biology, neuroscience, medicine, philosophy, literature. Now, having no further need to expand my scientific output or advance my academic standing, I have chosen to shift my approach. Instead of writing full-length articles for peer review, I now focus on recording and sharing original ideas, i.e., conceptual insights and hypotheses that I hope might inspire experimental work by researchers more capable than myself. I refer to these short pieces as nugae, a Latin word meaning “trifles”, “nuts” or “playful thoughts”. I invite you to use these ideas as you wish, in any way you find helpful. I ask only that you kindly cite my writings, which are accompanied by a DOI for proper referencing.

NUGAE - A GAUGE-THEORETIC MODEL OF THE HIERARCHICAL LEVELS OF HUMAN CONCEPTUAL KNOWLEDGE
Current theories of conceptual knowledge describe how humans organize ideas at multiple levels, from broad superordinate distinctions like living versus nonliving, to basic categories like animals and down to subordinate distinctions like dog, wolf or beagle. Modern cognitive science shows that people navigate these levels fluidly: they maintain stable category membership while flexibly reshaping the meaning of a concept depending on the situation. However, existing computational models often conflate these levels or fail to capture how meanings shift while the conceptual backbone is kept stable. Neural networks, for instance, can learn fine-grained similarities but struggle to express how context affects the interpretation of the very same item without altering its categorical identity. This reveals a missing mechanism in current models: a way to distinguish the stable structure of conceptual hierarchies from the flexible, contextual interpretation of specific concepts.
Our proposal addresses this gap by adapting a simplified version of gauge theory to conceptual representation. In physics, gauge theories describe systems whose internal configuration can change without altering their fundamental identity, like rotating a compass needle without moving the compass itself. Translating this intuition to cognition, we consider each concept as having two parts.
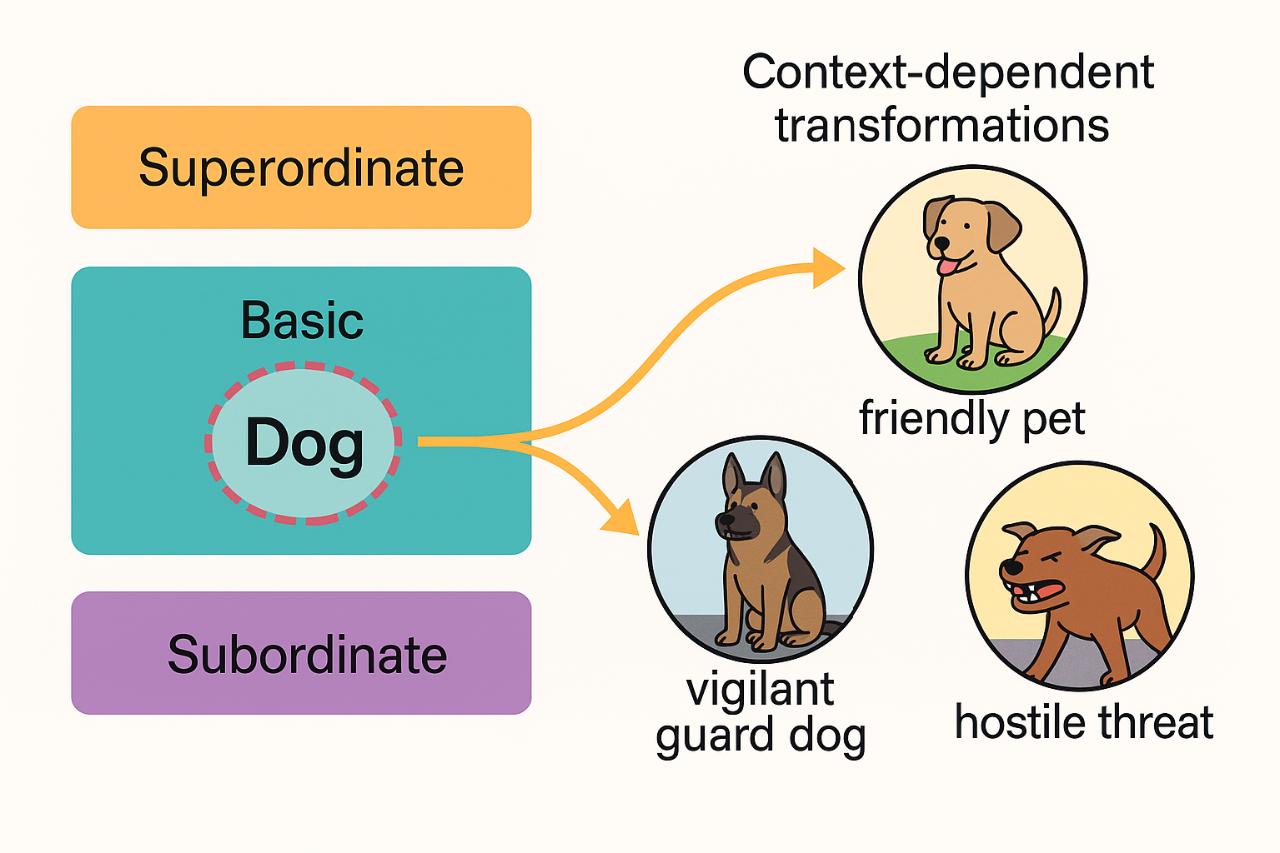
- The stable component, which situates it on the semantic hierarchy (dogs are animals, animals are living beings).
- The flexible component, made of microfeatures whose relevance changes with the context.
A dog provides a clear illustration. At the hierarchical level, it is always an animal, living and distinct from tools or plants; this position does not change. But the internal meaning shifts dramatically depending on the situation. When adopting a “family-home” frame, a dog is affectionate, familiar and safe. In a “security” frame, it becomes vigilant, strong and protective. Under a “danger” frame, the perceived threat is amplified. These shifts do not modify the concept’s place in the hierarchy; instead, they alter the internal feature weighting. Gauge transformations are used to formalize these context-driven changes. They reorganize the microfeatures without moving the concept from its semantic location, much like reorienting the compass needle without moving the compass.
In our framework, concepts lie on a manifold representing the hierarchical levels, while each concept carries a vector of flexible attributes attached like a small fiber. Contexts act as gauge fields: they apply structured transformations to the fibers, adjusting how the concept is internally represented. This grants a direct way to model how the meaning of a dog changes when one switches from thinking about pets, to thinking about dangerous animals, to thinking about working dogs, all while preserving the stability of the underlying hierarchy.
Compared with embedding-based approaches, our framework clearly separates identity from context, allowing formal control over what changes and what remains fixed. It also predicts structured patterns of semantic deformation: concepts within the same category should shift together under certain contextual frames, while remaining anchored to their hierarchical level.
This approach suggests several experimental tests. Behavioral similarity judgments should reveal systematic changes in perceived distances between items when the contextual frame is altered. Neuroimaging-based representational similarity analyses should show that coarse-level categories remain stable while fine-grained relations reorganize. Simulations should reveal predictable anisotropies, i.e., greater deformation along certain conceptual dimensions but not others.
Many potential applications can be suggested: modelling conceptual flexibility in language and reasoning, improving AI’s alignment with human conceptual structure, understanding semantic drift and framing effects and studying alterations of conceptual processing in clinical populations. Our approach invites further research into context-sensitive meaning, developmental changes in concept representation and cross-cultural variability in hierarchical organization.
QUOTE AS: Tozzi A. 2025. Nugae - a gauge-theoretic model of the hierarchical levels of human conceptual knowledge. DOI: 10.13140/RG.2.2.30644.10882
NUGAE - A PANDEMONIUM MODEL OF MORAL CHOICES
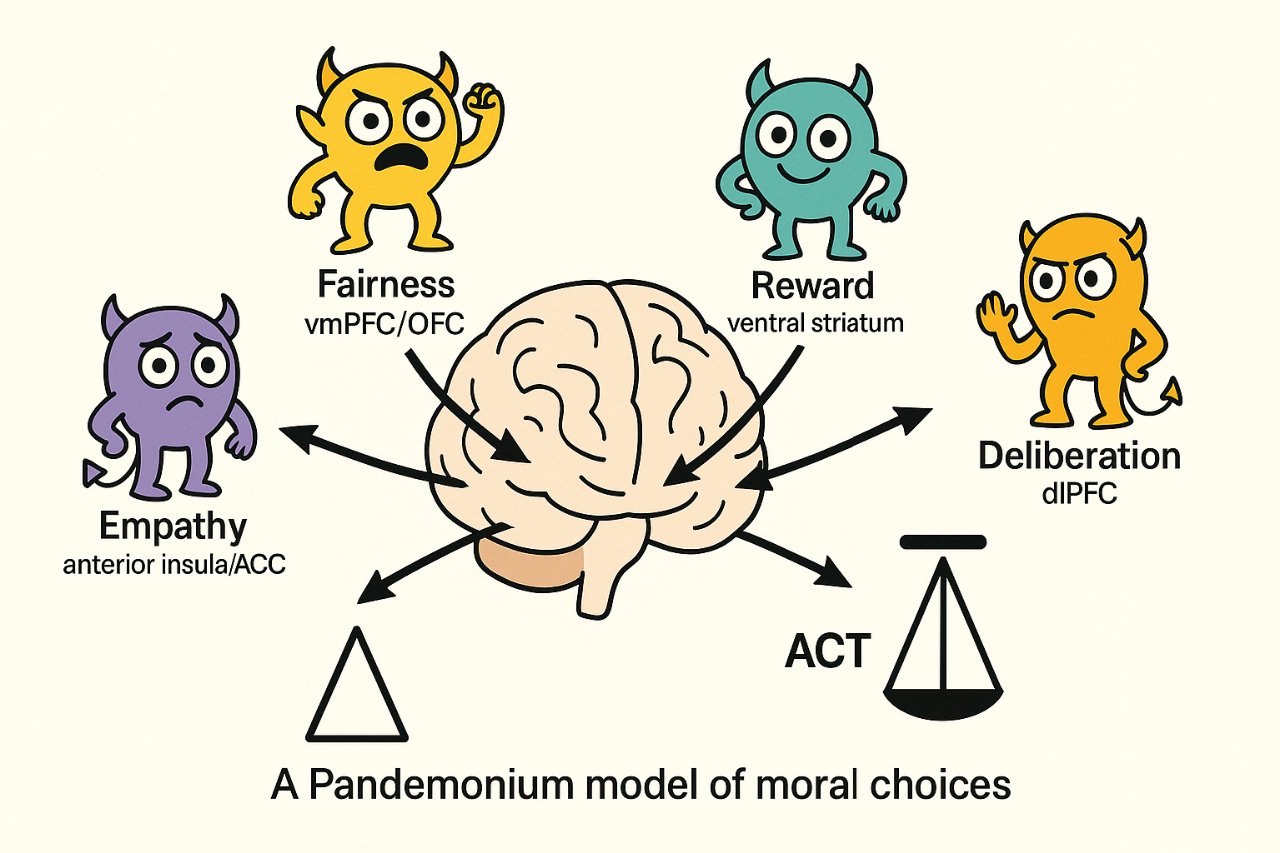
Empirical studies increasingly show that moral decision-making arises from distributed and partially independent neural systems whose activity patterns vary dynamically with context. Rather than assuming a central moral faculty or hierarchical control, we propose a Pandemonium model in which moral judgment results from the real-time competition and cooperation among multiple neural subnetworks. Each subnetwork operates as a computational demon, evaluating the same perceptual and contextual inputs under its own objective function. The outcome of this collective dynamic is not a predetermined rule, but rather an emergent equilibrium reflecting the transient dominance of specific evaluative dimensions. The paper contains some mathematical formulas that are difficult to reproduce in this context. Therefore, you can find here a Word version of the article
QUOTE AS: Tozzi A. Oct 2025. Nugae - A Pandemonium Model of Moral Choices. viXra:2510.0033
NUGAE - QUALIA MAPPED ONTO MULTIDIMENSIONAL MANIFOLDS OF POSSIBLE AND PROBABLE STATES
The study of qualia has long been caught between two poles: descriptive phenomenology, which stresses the richness and irreducibility of subjective experience and reductive science, which seeks to capture experience through objective correlates such as wavelengths, decibels or neural activations. Both approaches face limits: what is lacking is a framework preserving the open-ended differentiability of qualia while also allowing for tractable representation.
We suggest treating qualia as points and trajectories on a multidimensional manifold, in which every potential axis corresponds to a possible distinction that experience could make salient, e.g., hue, brightness, texture, emotional valence, bodily resonance, temporal unfolding and so forth. Rather than collapsing experience into a single category, we aim to expand it into a geometric space where fine-grained differences can be located and compared.
Crucially, we distinguish between possible distinctions, i.e., the full phase space of the discriminations that the perceptual and cognitive system could in principle support, and probable distinctions, which are the actually realized, ecologically weighted and historically constrained differentiations employed by subjects. The manifold provides the scaffold, while probability distributions assign density to its regions, indicating where distinctions are more likely to occur. In this way, qualia are represented not as fixed states but as probabilistically weighted trajectories through a high-dimensional experiential landscape. Our formulation provides advantages over existing models of perceptual or affective space, which either focus narrowly on one modality or assume fixed coordinate systems.
Experimental tests could involve psychophysical paradigms where subjects discriminate finely graded stimuli, combined with neural population analyses to assess manifold structures in brain activity. Topological data analysis, diffusion maps and embedding techniques could approximate the geometry of possible distinctions, while behavioral frequencies and neural likelihoods could estimate the distribution of probable distinctions. Testable predictions include the emergence of invariant attractors corresponding to recurrent qualia categories and measurable differences between the geometry of possible perceptual spaces and the densities of probable usage.
Our approach may lead to new strategies for interpreting perceptual plasticity and for diagnosing disorders where the weighting of distinctions is altered. Still, it may inspire artificial intelligence architectures balancing open-ended representational capacity with probabilistic efficiency. In philosophy, the intractability of qualia could be reframed not as a dead-end, but rather as a structured manifold awaiting exploration. Future research could extend our framework to collective qualia spaces, intersubjective resonance, multidimensional cues and cross-species comparison, deepening our grasp of how worlds of experience are constituted.
It would also be possible to use the same language to describe what a bat, a lion or a human perceives.
QUOTE AS: Tozzi A. 2025. Nugae -qualia mapped onto multidimensional manifolds of possible and probable states. DOI: 10.13140/RG.2.2.23426.75205

NUGAE – IS POETRY THE MIND’S ORIGINAL LANGUAGE?
Which is the language used by the brain to think? the language of thought remains debated. Hypotheses include Mentalese (Fodor), a symbolic inner code resembling logic; neural codes, where meaning is carried by firing patterns and oscillations; predictive processing, where the brain constantly models and updates expectations; embodied cognition, grounding thought in sensorimotor patterns. Here we explore the possibility that humans (and animals) engage in a poetic form of mental language when they think.
Poetic language is characterized by a set of features that make it different from ordinary discourse. It relies on metaphor and analogy, where abstract concepts are expressed through vivid cross-domain mappings that make complex ideas more graspable. Repetition and parallelism reinforce memory and create a sense of cohesion, while imagery and symbolism compress layered meanings into concise and evocative forms. Unlike strictly literal language, poetic language embraces ambiguity and polysemy, sustaining multiple interpretations simultaneously and inviting flexible thought. Its power is heightened through sound play, including rhyme, meter, alliteration and assonance, which exploit the brain’s sensitivity to auditory regularities and contribute to structure expression and guide attention. Poetic language often fosters emotional resonance and collective synchrony, linking individual cognition to shared experience.
HUMANS. Research across cognitive science, neuroscience and literary theory converges on the idea that poetic structures are correlated with human cognition. Cognitive linguistics demonstrated that metaphors are not ornaments but the scaffolding of abstract thought (Lakoff & Johnson). The field of cognitive poetics links features such as rhythm, sound play and ambiguity to mental processing (Tsur, Steen, Gavins), while Arthur Jacobs’ neurocognitive poetics model examines how literary reading recruits distinct neural circuits integrating emotion and cognition. Neuroimaging studies reveal that poetry activates broader cortical and limbic networks than prose, engaging memory, imagery and affective systems. Research on rhythm and prosody highlights their role in prediction, attention and learning, while work by Wassiliwizky and others shows that poetic language elicits measurable emotional and physiological responses. Holyoak’s analysis of metaphor underscores its dual role in science and art. It is noteworthy that neuroscience emphasizes the very rhythmic and oscillatory nature of language processing in the brain. Evolutionary anthropology points to the ubiquity of chant, song and poetic parallelism in early human societies, suggesting that poetic structures may be far more ancient and essential than the prose-like logic that later came to dominate science and philosophy.
Collectively, multidisciplinary studies suggest that poetry-like structures pervade cognition, though no unified model yet exists. Indeed, current theories of cognition emphasize mechanisms such as predictive processing, working memory and symbolic representation, yet they often treat poetic expression as a secondary cultural artifact rather than a central cognitive strategy.
We propose that humans do not merely produce poetry as an art form, but actually think poetically by default. This implies that metaphor, rhythm, repetition, imagery, ambiguity and sound play are not stylistic additions, but rather cognitive necessities embedded in neural architecture. We also suggest neural correlelates for poetic features (see figure). Metaphor may correspond to cross-domain conceptual mapping in the parietal and prefrontal cortices, rhythm to neural oscillations that synchronize communication across brain regions, imagery to hippocampal-visual interactions that compress complexity into symbolic representations. Ambiguity may allow the prefrontal cortex to sustain multiple possible interpretations before collapsing into choice, while sound patterns exploit auditory cortex sensitivity to phonological regularities. Collective chanting may reflect the synchronizing effect of rhythmic speech on group neural activity and hormone release, fostering cohesion and memory transmission.
Compared with existing paradigms that approach language as symbolic code or information processing, our framework emphasizes the brain’s reliance on rhythmic, metaphorical and emotionally resonant structures: rather than seeing poetry as a cultural luxury, we see it as a natural cognitive technology.
Experimental tests of our hypothesis could involve neuroimaging studies that compare the processing of poetic versus prosaic material, focusing on whether poetic features enhance predictive coding efficiency, memory consolidation and neural synchrony. Testable hypotheses include that metaphor-rich texts activate broader cortical networks than literal texts, rhythmic language enhances theta–gamma coupling and ambiguous verses sustain prefrontal activity longer than unambiguous statements.
Applications range from education, where poetic structures could enhance learning, to therapy, where rhythm and metaphor may regulate emotion and foster resilience. Future research could expand to artificial intelligence, where poetic models might inspire new architectures capable of flexible, image-rich and ambiguous reasoning, offering machines not only the ability to compute but also to think in patterns closer to the human mind.
ANIMALS. We propose that animals, too, may think and communicate in proto-poetic forms. Indeed, honeybee’s dance, birdsong, whale song, primate calls, ritualized displays such as the peacock’s tail, show rhythmic structuring, repetition with variation and symbolic signals, suggesting that poetic features may be part of a broader evolutionary substrate. Unlike current approaches treating animal communication as rigid code-like signals, we emphasize patterned, emotionally charged and flexible structures enabling memory, prediction and social cohesion.
Experimental tests could compare responses to rhythmic versus arrhythmic signals, assess sensitivity to repetition and variation and apply entropy-based analyses to detect metaphor-like compression in call sequences. Testable previsions include that animals preferentially attend to rhythmic or repeated patterns, show neural entrainment to structured signals and use ambiguous vocalizations to maintain flexible responses.
Applications include more precise interpretation of animal signals by revealing patterned, metaphor-like structures in vocalizations, gestures or ritualized displays that might otherwise be overlooked. Future research could establish quantitative metrics of “poeticity” to systematically compare human and animal cognition, explore evolutionary continuities and test whether patterned, metaphor-like signalling constitutes a shared substrate of intelligent communication across species.
QUOTE AS: Tozzi A. 2025. Nugae - is poetry the mind's original language?. DOI: 10.13140/RG.2.2.16891.17445

NUGAE - NEURAL RIGID DESIGNATORS: HOW THE BRAIN RECOGNIZES YOUR WIFE, NO MATTER THE CHANGES
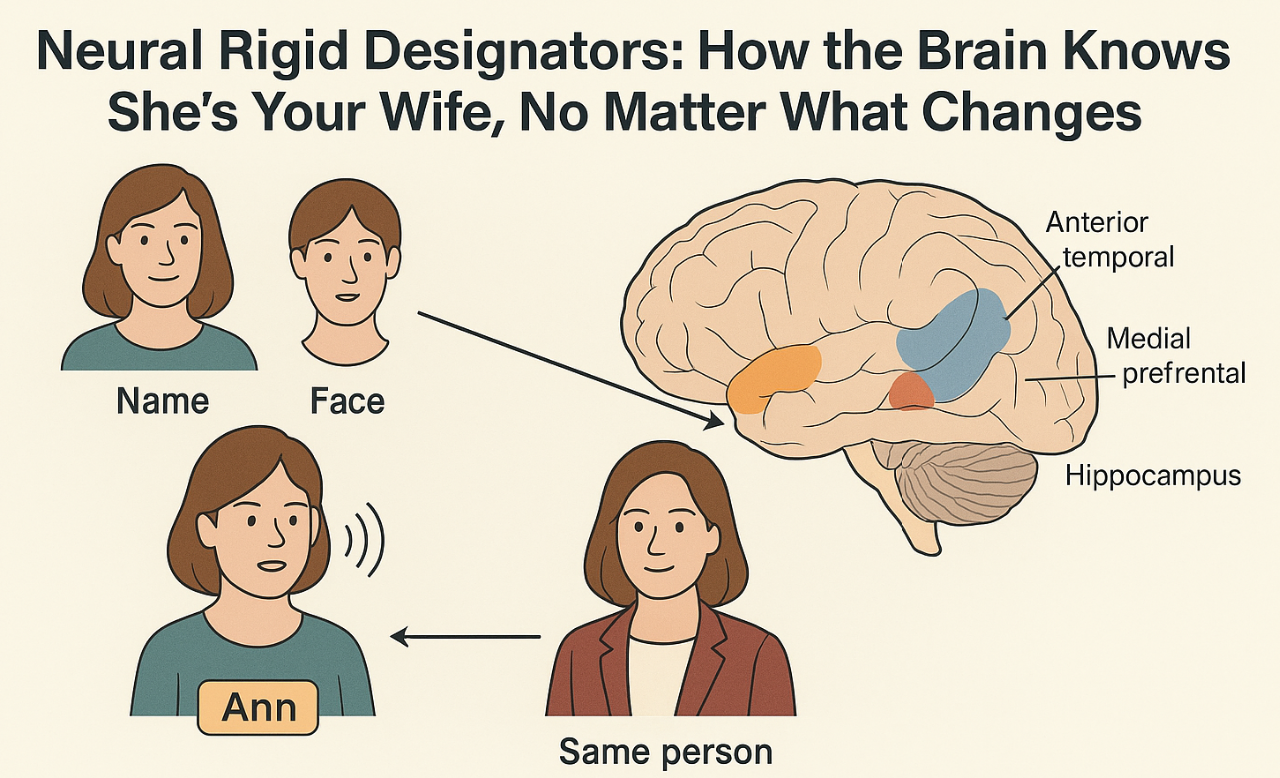
The brain recognizes the same person or object across large changes in appearance, viewpoint, sound and context. Current neuroscientific accounts usually treat identity representations as rich bundles of features or as similarity patterns in a high-dimensional space. What is still lacking is a bridge between the neural code that fixes an entity’s unique identity (its “who”) and the distinct codes that capture its “what,” namely the features, roles and properties that can change over time or across contexts.
Building on Kripke’s philosophical account, we propose that identity coding in the brain can be understood as a neural rigid designator, i.e., an internal mechanism consistently referring to the same entity across all situations. It may operate as a stable pointer established at the time of learning and reactivated whenever that entity is encountered, irrespective of changes in its descriptive features. Functionally, it may behave like a label attached to a latent binding slot that persists across time and contexts. The hippocampus may anchor it to episodic traces, the anterior temporal hub may connect it to semantic attributes and medial prefrontal circuits may regulate its selection according to context and goals.
Proper names, faces, voices, and symbolic cues can all trigger the same underlying code, which then acts as a key, granting access to information about the individual stored elsewhere in the brain and adapted to the current task. Crucially, this code might remain stable even when surface features (like lighting, hairstyle, social role, etc) change, much like a permanent entry in a mental index that ensures recognition regardless of appearance or circumstance.
Compared with feature-centric or similarity-only approaches, our neural rigid designator could separate reference from description; explain robust cross-modal linking between names, faces and voices; account for dissociations where people lose proper names while retaining descriptive knowledge; support long-range generalization in new settings.
Methods to test this account could employ representational similarity and encoding models with fMRI and MEG to track identity codes across variations in features and tasks; use cross-decoding to link names with faces or voices; measure repetition suppression as evidence of code reuse; apply causal interference through TMS or intracranial stimulation targeting anterior temporal and hippocampal regions.
Testable hypotheses can be drawn from our hypothesis: identity codes might remain linearly decodable across major changes in surface features; disrupting anterior temporal or hippocampal function might weaken their persistence, while leaving property recognition intact; proper name cues might reinstate the same code activated by perceptual cues; counterfactual alterations of features might not compromise their stability.
Theoretical applications include: biomarkers for prosopagnosia and semantic dementia; memory-supportive brain computer interfaces; human-robot systems keeping person identity consistent across modalities; AI architectures with entity pointers for co-reference and lifelong learning. Future research could map developmental emergence, plasticity after injury, interference limits as the number of identities grows and extension from concrete individuals to abstract entities like institutions or theories.
QUOTE AS: Tozzi A. 2025. Nugae - Neural rigid designators: how the brain recognizes your wife, no matter the changes
doi: 10.13140/rg.2.2.34051.62249
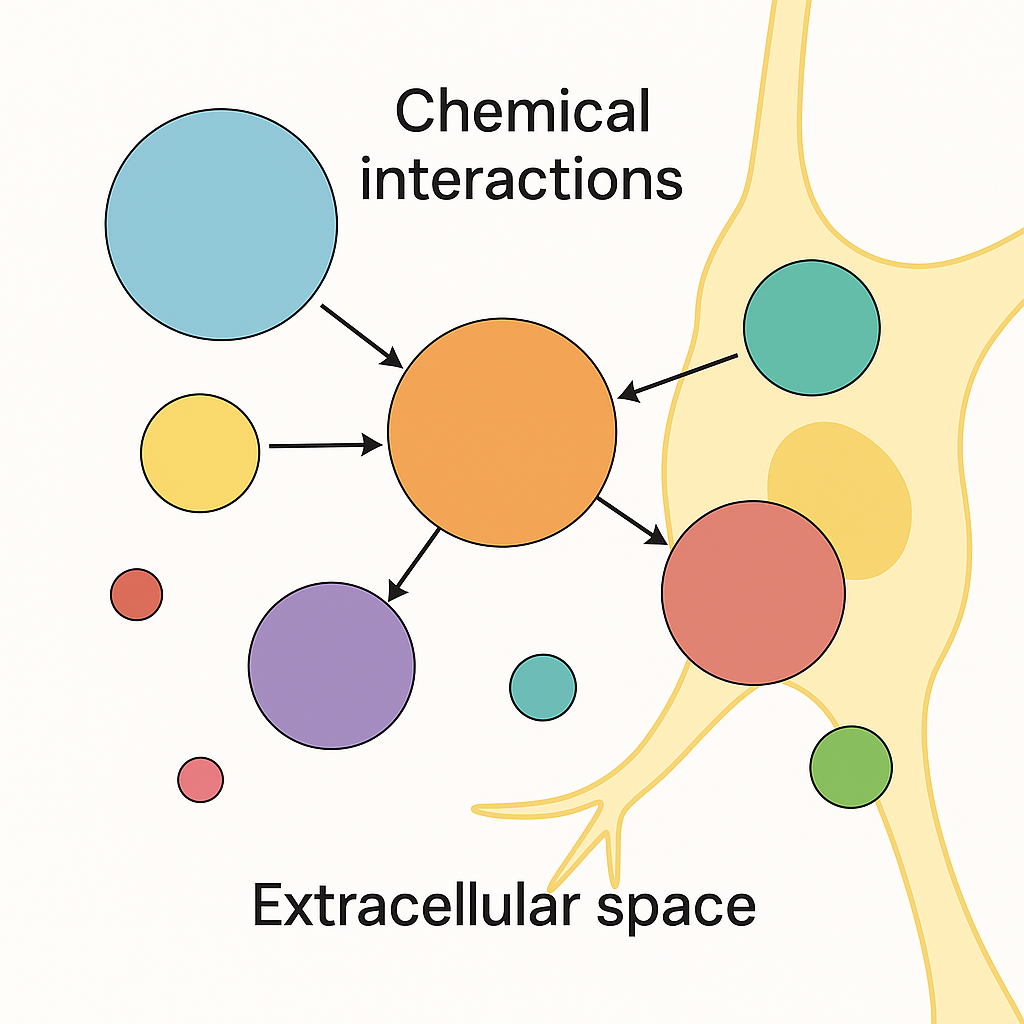
NUGAE - TOWARD A CHEMICAL CONNECTOME OF THE BRAIN’S EXTRACELLULAR SPACE
In neurobiological models, chemical signaling is usually described as a set of linear or localized interactions between neurotransmitters and their receptors. While these approaches capture aspects of fast synaptic transmission and receptor binding specificity, they fail to account for the dynamic complexity of the extracellular chemical environment in the brain. This includes the coexistence and mutual modulation of neurotransmitters, neuromodulators, hormones, peptides, neurosteroids and ions within the perineuronal space. Still, connectome frameworks describe physical or functional connectivity but overlook the biochemical layer where molecules diffuse, interact nonlinearly and influence neural dynamics beyond the synaptic borders.
We propose a framework to conceptualize chemical signaling layer as a high-order network composed of interacting molecular species. Each chemical messenger may be modeled as a node and the edges may represent regulatory, modulatory or competitive influences (including, e.g., enzymatic degradation, transporter competition, receptor cross-talk and co-release). Higher-order connections may capture convergence on common second messenger systems, feedback loops within endocrine-neural circuits and shared structural modulators like the extracellular matrix. A chemical connectome may account for time-dependent features like circadian hormone release, activity-dependent diffusion and stress-responsive neurochemical shifts, all embedded in a spatially defined topology reflecting region-specific receptor expression and extracellular gradients. Our approach could represent emergent properties of brain chemistry such as redundancy, buffering, antagonism, synergy and temporal gating, phenomena poorly captured by pairwise or purely anatomical connectomes.
To test experimentally our hypothesis, one could employ multi-modal molecular imaging, genetically encoded neurotransmitter sensors and real-time microdialysis in combination with optogenetics or pharmacological perturbation to monitor how changes in one messenger affect the spatial distribution and activity of others. Testable hypotheses include whether elevation of one transmitter (e.g., dopamine) dynamically alters extracellular half-lives or receptor availability of others (e.g., serotonin or glutamate) or whether co-released peptides (e.g., NPY) extend or buffer classical neurotransmission. Applications range from simulating drug interactions to elucidating neurochemical mechanisms of psychiatric disorders, in which dysfunction may arise from widespread network-level imbalances rather than isolated receptor abnormalities. In neuroengineering, our approach could inform artificial neuromodulatory control in brain-computer interfaces.
Future research may explore how high-order chemical networks interface with structural and functional networks in the brain, forming a multi-layered chemical connectome that encompasses molecular interactions. Practical examples include serotonin inhibiting glutamate release presynaptically, dopamine and serotonin competing for monoamine oxidase degradation or neurosteroids like allopregnanolone enhancing GABAergic inhibition via allosteric modulation.
Overall, the extracellular space may stand for a computational milieu where chemical signals do not simply coexist but actively constrain and enable one another’s functions.
QUOTE AS: Tozzi A. 2025. Nugae -toward a chemical connectome of the brain's extracellular space. DOI: 10.13140/RG.2.2.22042.15046.
NUGAE - MODELS OF NEURAL REPRESENTATION AND BINDING BASED ON PRIME NUMBERS
Neuroscience has long sought efficient and unambiguous coding schemes to assess how neural systems represent, bind and multiplex information. Various current models rely on spike timing, rate codes, oscillatory phase coding and population vector approaches, which successfully capture several aspects of neural computation but often struggle with the huge amount of possibilities, the binding problem, etc.
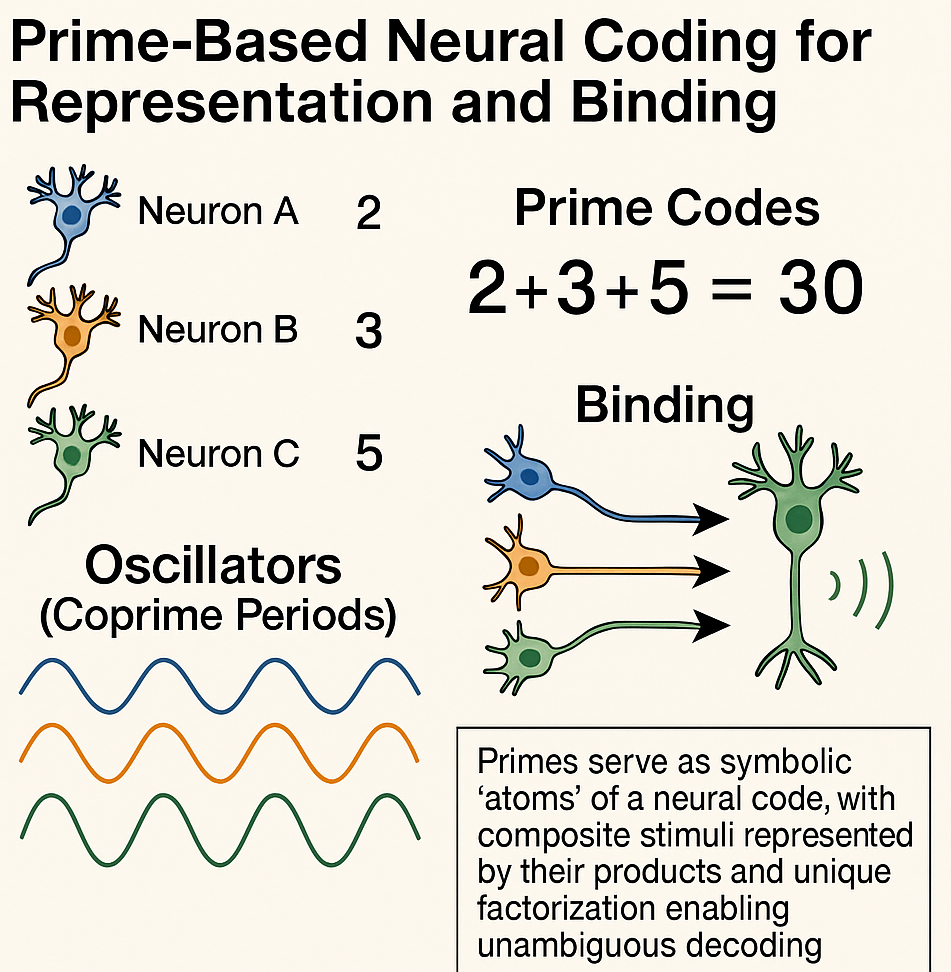
Prime number theory may provide a mathematical alternative: primes could be used as indivisible symbolic atoms for neural codes, leveraging the unique factorization theorem to guarantee that any combination of features can be represented by a single composite number and decoded without ambiguity. Each neuron, feature or informational unit could be assigned a prime: simultaneous activation corresponds to multiplication of their assigned primes, while decoding simply requires factorization. This leads to a biologically plausible implementation by mapping primes to residues in oscillators whose cycle lengths are coprime. This would enable Chinese Remainder Theorem–based phase code that the brain could implement through well-known oscillatory microcircuits like theta–gamma coupling (see the CODA for more technical details).
Compared to existing approaches, our prime numbers approach to brain computations provides built-in noise robustness, since small perturbations in spike timing or phase cannot alter the factorization unless the disturbance is extreme. The model scales efficiently, as a small set of primes can encode vast combinatorial repertoires and allows multiplexing of independent streams without interference.
Experimentally, the model could be tested by training spiking networks or neuromorphic circuits with oscillators set to coprime periods and teaching them to represent feature combinations, then measuring decoding accuracy and resilience to noise. In vivo, one could look for task-dependent oscillatory components with coprime periods and phase signatures corresponding to learned feature conjunctions. Testable hypotheses include: during tasks requiring complex binding, narrowband oscillations will emerge at coprime-related frequencies; lesions to specific oscillators will cause predictable “arithmetic” decoding errors; specific combinations of features will yield rare cross-frequency alignment events corresponding to the least common multiple of their assigned periods.
Potential applications span from improving artificial neural networks’ ability to bind and unbind features, to brain–computer interface encoding schemes, to error-resilient communication protocols inspired by neural computation. Future research could explore adaptive prime assignment through synaptic plasticity, hybrid architectures combining number-theoretic representations with probabilistic inference and integration of prime-based codes with rate and temporal codes.
CODA. Biologically-plausible neural architectures exploiting prime factorization could be built by assigning each informational feature (e.g., a neuron, or a microcolumn, etc.) a unique prime-based code. A small set of neural oscillators, each with a cycle length corresponding to a pairwise coprime or prime number, would serve as a temporal scaffold. When a feature is activated, it induces precise phase shifts in each oscillator according to its assigned residues. When multiple features are active, these phase shifts sum, performing multiplicative binding through the Chinese Remainder Theorem. Phase-selective neurons would then decode the residue pattern from the oscillators and an associative microcircuit could reconstruct the original feature set. Rare alignments of all oscillators, corresponding to the least common multiple of their periods, would signal specific conjunctions, creating a natural detection mechanism for complex bindings. Learning of the prime-residue mapping could occur through spike-timing-dependent plasticity, while neuromodulatory signals would control whether the network operates in a write or read mode. Overall, this theoretical architecture would provide a compact, noise-resilient and combinatorially rich coding system, potentially allowing the brain to represent vast numbers of feature combinations with minimal resources.
QUOTE AS: Tozzi A. 2025. Nugae -models of neural representation and binding based on prime numbers. DOI: 10.13140/RG.2.2.32082.26562
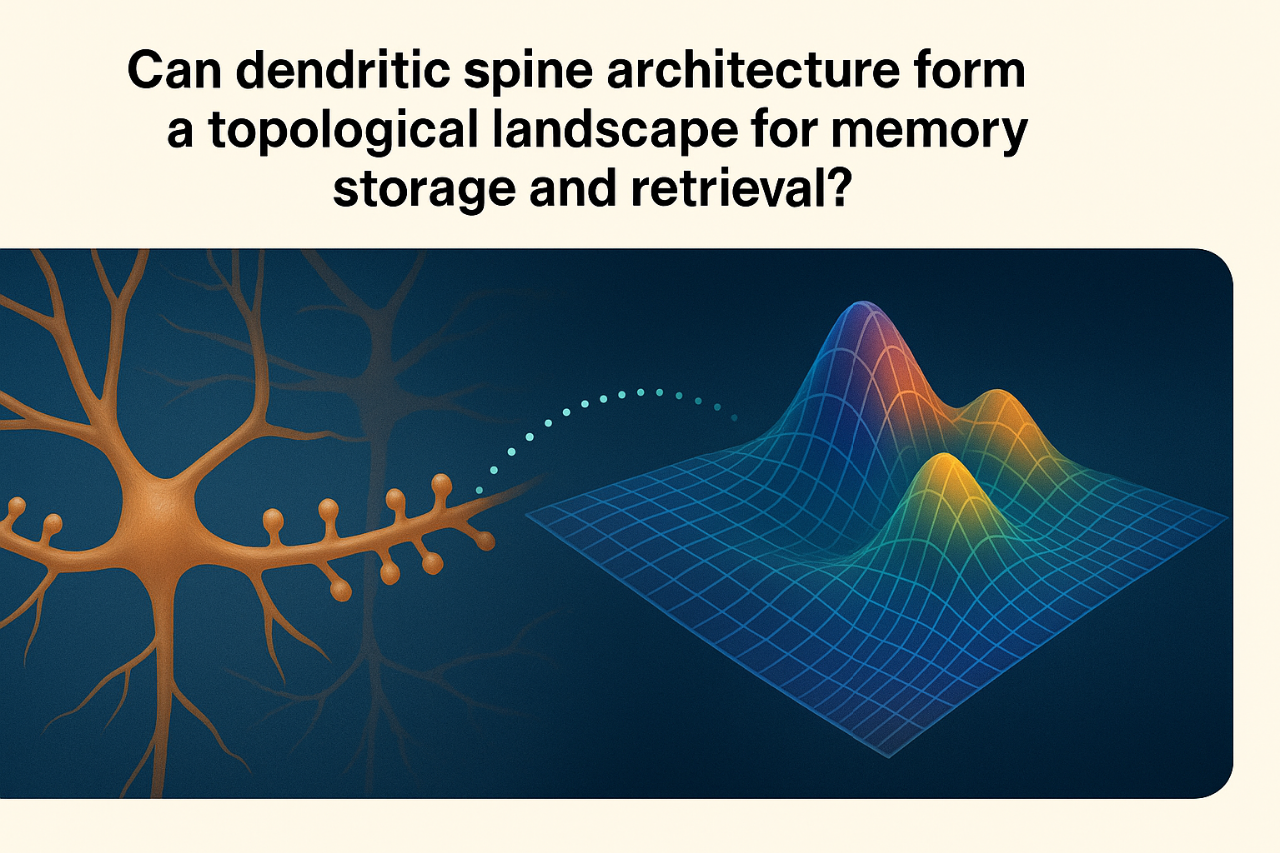
COULD DENDRITIC SPINE ARCHITECTURE SHAPE A TOPOLOGICAL LANDSCAPE FOR MEMORY STORAGE AND RETRIEVAL?
We hypothesize that memory may emerge from the spatial arrangement of countless dendritic spines across neurons, giving rise to a topological landscape that supports distributed, resilient and overlapping encoding/retrieval across neural networks.
Memory storage involves synaptic plasticity, i.e., changes in synaptic strength and number, driven largely by the remodelling of dendritic spines. Long-Term Potentiation, AMPA receptor trafficking and spine clustering are core mechanisms in this model. Nevertheless, these explanations remain local and linear, assuming memory is encoded at specific sites and strengthened through repetition. While useful for describing short-term and mechanistic changes, they fall short in accounting for how memory is distributed, redundant, resilient and capable of flexible association and recall..
We hypothesize that memory is not stored at isolated synapses or confined to discrete neural units. We propose that the spatial arrangement of countless dendritic spines across neurons gives rise to a high-dimensional, transient topological manifold, in which each spine represents a spatial coordinate within that structure. Dendritic spines may serve not merely as passive recipients of synaptic input, but as active, spatially positioned nodes in a continuously evolving encoding landscape. Their patterns of spatial proximity, morphology and coordinated timing of activation may form a flexible field through which memory traces emerge, interact and persist.
Our framework suggests that memory traces may emerge as diffuse, resilient network-wide constructs, interwoven with other traces. One key implication is that even partial patterns (subsets of spine clusters or activity waves) could be useful to reconstruct the full or approximate content of a memory. This aligns with observations in cognitive neuroscience, where memory recall can be triggered by fragmentary cues or partial reactivation. Also, it may explain how overlapping memories can coexist without overwriting one another, and how degraded or noisy signals can still yield meaningful retrieval. Still, our topological account of the dendritic spine manifold defines memory as a spatially organized, emergent process, potentially explaining how memories can be redundant, overlapping, associatively linked, and both resilient and adaptable over time.
To experimentally investigate our hypothesis, one could use two-photon imaging to track spine growth, decay and clustering during learning tasks. Cell tagging with optogenetics would allow researchers to link specific spine patterns to memory traces. High-resolution 3D electron microscopy reconstructions could analyze the geometry of spine distributions, while computational modeling might simulate spine interference fields using wave-based algorithms.
To uncover hidden geometric patterns in memory-related spine arrangements across time and neuronal populations, topological techniques such as persistent homology, simplicial complexes and manifold learning (e.g., UMAP, t-SNE) can be applied. These methods may enable analysis of the shape, connectivity, clustering and high-dimensional structure of evolving dendritic spine distributions.
In artificial intelligence, our model may inspire biologically grounded memory architectures that rely on spatial pattern encoding rather than rigid storage units. In neuroscience, it may reframe neurodegeneration not as the loss of individual synapses, but as the disruption of coherent spatial patterns, potentially enabling earlier and more targeted diagnostics. Clinically, it opens the door to interventions for trauma and memory disorders by aiming to restore or reinforce disrupted spatial memory structures.
QUOTE AS: Tozzi A. 2025. Nugae -Could Dendritic Spine Architecture Shape A Topological Landscape For Memory Storage And Retrieval? DOI: 10.13140/RG.2.2.34279.36002